如果您无法下载资料,请参考说明:
1、部分资料下载需要金币,请确保您的账户上有足够的金币
2、已购买过的文档,再次下载不重复扣费
3、资料包下载后请先用软件解压,在使用对应软件打开
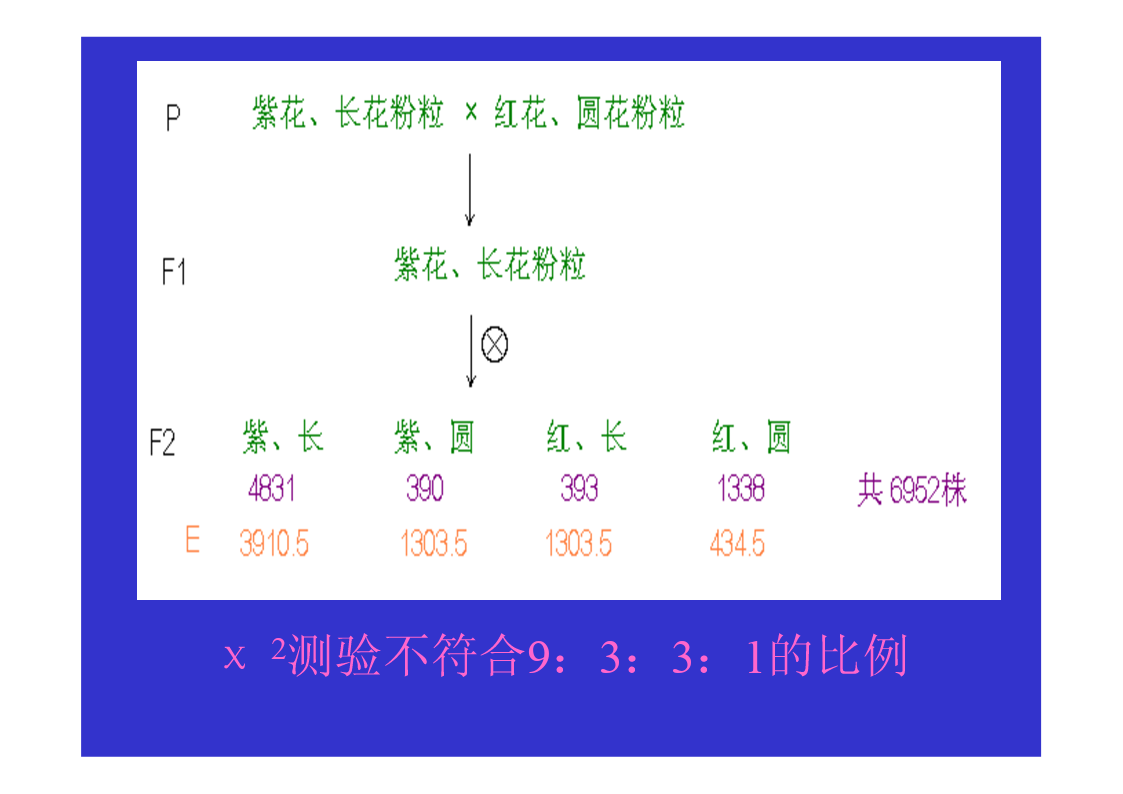
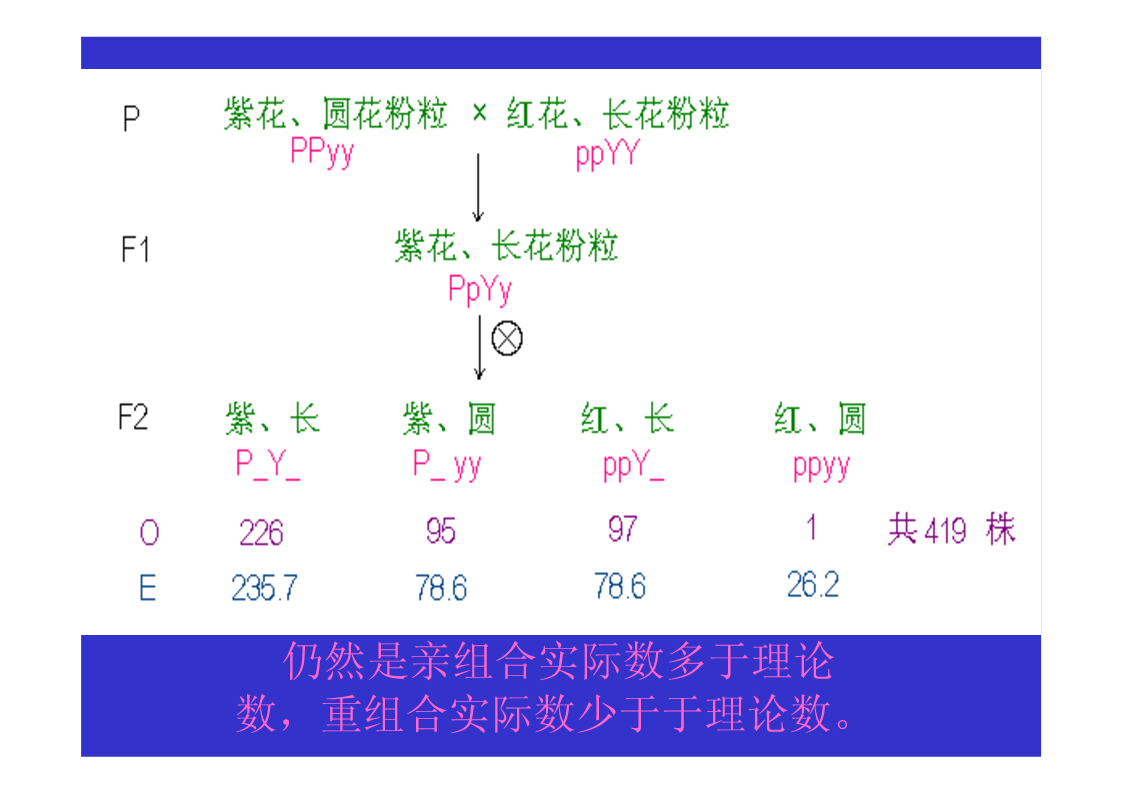
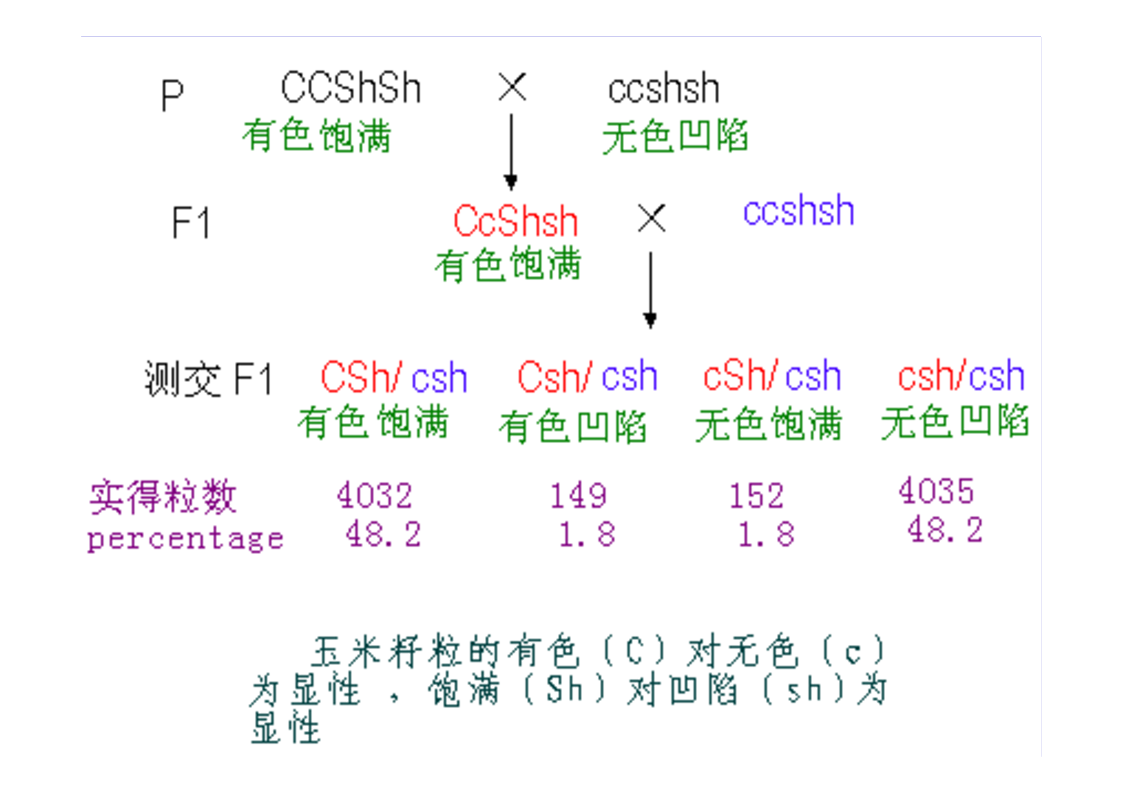
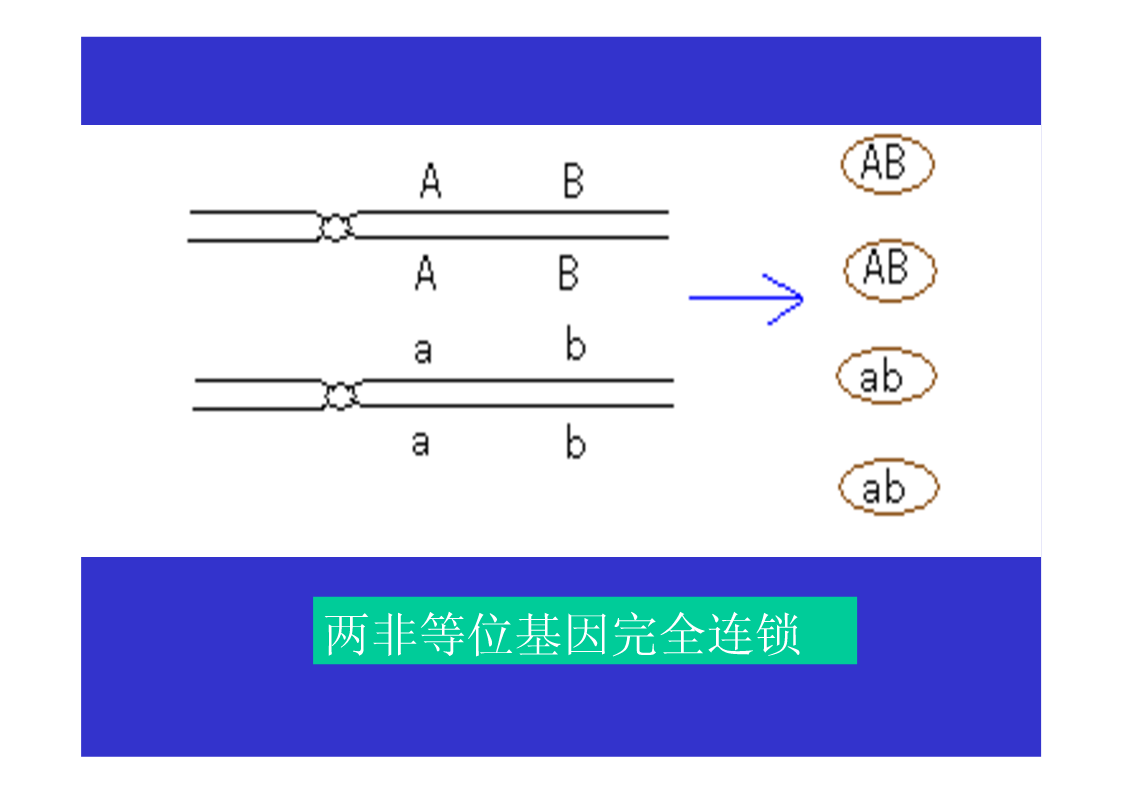
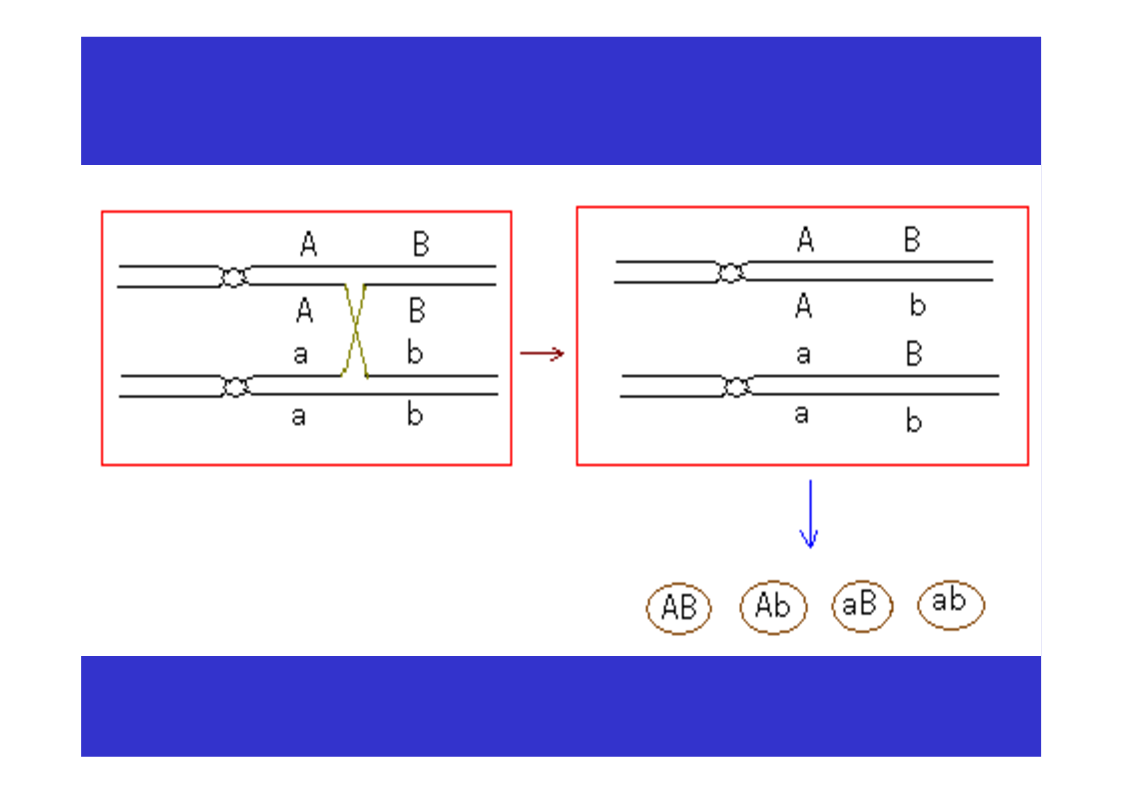
第三节连锁与交换一、连锁现象的发现Bateson等1906,香豌豆杂交试验χ2测验不符合9:3:3:1的比例仍然是亲组合实际数多于理论数,重组合实际数少于于理论数。原来为同一亲本所具有的两个性状,在后代常常有连锁在一起遗传的倾向。二、连锁遗传的本质F1形成的四种配子数不等三、连锁互换的细胞学基础自由组合的细胞学基础AaBb→AB,Ab,Ab,ab四种配子基因与染色体的平行现象两非等位基因完全连锁若F1在形成配子时,100%的性母细胞在进行减数分裂时,在连锁的基因A、B座位之间,都发生了一次同源染色体非姊妹染色单体的交换,则各种配子比例情况…若80%的性母细胞…,则配子比例情况…若7.2%的性母细胞…,则配子比例情况……综合练习题两个果蝇突变品系,一个是黄眼,另一个是鲜红眼。用黄眼雌性与鲜红眼雄性杂交,F1代有251个野生型雌性,248个黄眼雄性。F1中的雄性与雌性杂交,得到以下F2代表现型:260个野生型雌性,253个黄眼雌性,77个野生型雄性,179个黄眼雄性,183个鲜红眼雄性,80个棕眼雄性。用图示法解释上述结果。四、基因定位(一)连锁群连锁群(linkagegroup):位于同一染色体上的所有基因,构成了一个连锁群。基因定位:确定基因在染色体上的位置和距离,叫做基因定位。(二)基因定位的方法遗传作图(geneticmapping):应用动植物杂交技术、人类家系分析、、链孢霉顺序四分子分析、噬菌体复感染等,来构建基因图谱。遗传作图的基础都主要是通过连锁分析计算重组率。同源重组→同源染色体的非姊妹染色单体之间,在减数分裂过程中,发生交叉和交换,导致连锁基因重组的现象。简单地说,就是连锁基因的互换所导致的遗传重组。五、重组率的计算与遗传作图(一)重组率的基本定义重组型配子数RF=×100%总配子数RF:RecombinantFrequency重组率的最小值为0。重组率的最大值为50%。RF取值范围在0到50%之间。(二)影响重组率大小的因素1.两连锁基因之间的距离:在一定范围内RF与基因间距离成正比,所以用重组率代表基因之间的距离,重组率去掉%即为遗传图距。(当基因间距离增大时,不再与RF成正比,此时可利用2.特定基因的突变;Haldane作图函数校正。)3.环境条件例如温度等;4.性别:雄果蝇和雌蚕通常不发生交换(霍尔丹定律)。(三)测交分析计算重组率1.两点测验F1测交后代基因型种类和比例即反映了双杂合体所产生的配子类型和比例,据此可直接计算RF(=2Y)。如果要测定3个连锁基因的顺序与距离,则需要进行3次两点测验,共进行3次杂交、和3次测交!!!自交法测定重组率2.三点测验2.1试验原理通过一次杂交、一次测交,同时确定三个连锁基因的顺序和距离。测交后代的8种基因型,按它们的产生途径以及所占比例,可分为4大类:在上表中①根据双交换的基因型类型,确定三个连锁基因的相对顺序;2.2双交换与并发系数①理论双交换值:等于两边两个重组值的乘积。如上图中:理论双交换值=RF(a-b)×RF(b-c);②干涉(interference):一个单交换发生后,在它临近再发生第二个单交换的机会就会减少,这种现象称为干涉或干扰。③并发系数(coefficientofcoincidence)干涉的程度用并发系数(或称符合系数)表示:并发系数C=实际双交换值理论双交换值;C通常变化在0~1之间。等于1表示无干涉;等于0表示干涉完全。但C也可能大于1,这种现象称为负干涉。2.3例题讲解用T4病毒的两个品系感染大肠杆菌细胞,一个品系是小噬菌斑(m)、快速溶菌(r)和噬菌斑混浊(tu)突变型;另一品系对这3个标记皆为野生型(+++)。把上述感染的溶菌产物涂平板,结果如图。a.测定m,r和tu三个基因的连锁顺序和距离?b.这个杂交的并发系数是多少,它意味着什么?解题步骤:①数据与基因型归类②比较亲型与双交换基因型,确定基因顺序。→r在中间位置,顺序为m-r-tu。③求基因之间的两两重组值。RF(m-r)=×100%=12.84%;RF(r-tu)=×100%=20.81%;RF(m-tu)=×100%=27.19%.④求并发系数C理论双交换值=RF(m-r)×RF(m-tu)=12.84%×20.81%=2.672%;实际双交换值=(162+172)