如果您无法下载资料,请参考说明:
1、部分资料下载需要金币,请确保您的账户上有足够的金币
2、已购买过的文档,再次下载不重复扣费
3、资料包下载后请先用软件解压,在使用对应软件打开
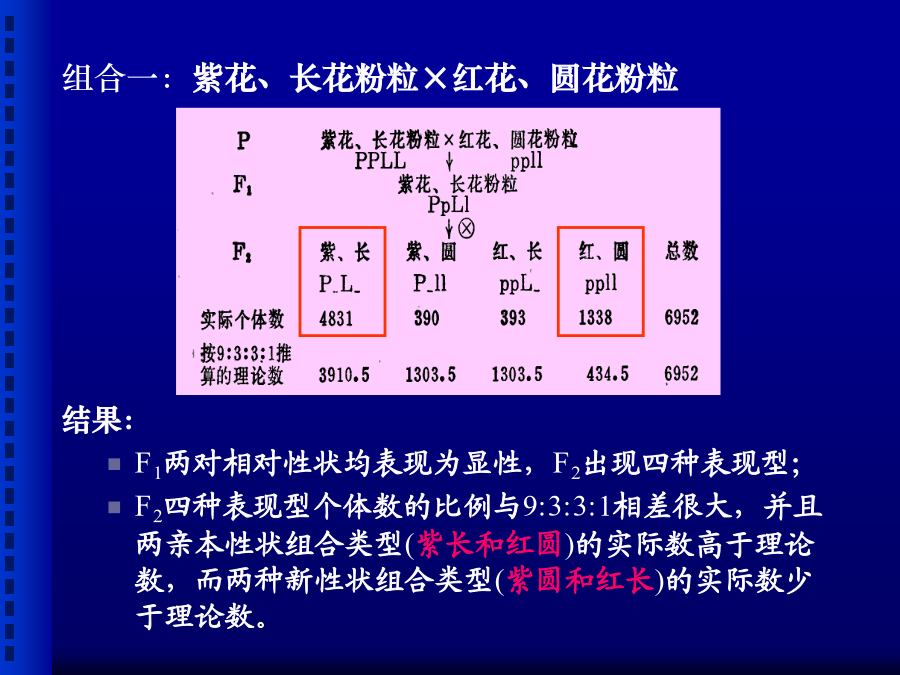
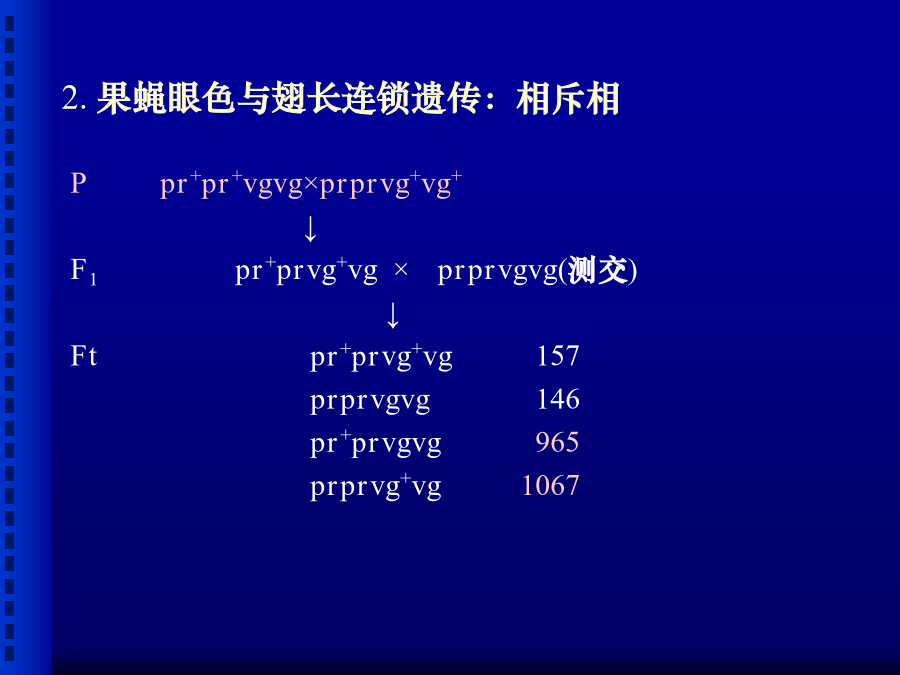
第五章连锁遗传和性连锁1900年孟德尔遗传规律被重新发现以后,许多学者以更多的动、植物为材料进行杂交试验,大量的杂交试验证实了孟德尔的遗传规律,但也发现有些多对性状的杂交实验结果并不符合独立遗传规律。摩尔根(1910)以果蝇为试验材料开展了深入细致的研究,最后确认所谓不符合独立遗传规律的一些例证,实际上属于另一类遗传,即连锁(linkage)遗传。摩尔根提出了遗传学中的第三个遗传规律—连锁遗传规律以及连锁与交换的遗传机理,并创立基因论(theoryofthegene)。摩尔根(T.H.Morgan,1866-1945)一、连锁组合一:紫花、长花粉粒×红花、圆花粉粒组合二:紫花、圆花粉粒×红花、长花粉粒连锁遗传现象:杂交试验中,原来为同一亲本所具有的两个性状在F2中不符合独立分配规律,而常有连在一起遗传的倾向,这种现象叫做连锁(linkage)遗传现象。相引相(couplingphase)与相斥相(repulsionphase)把甲乙两个显性性状连系在一起遗传,而甲乙两个隐性性状连系在一起遗传的杂交组合,称为相引相或相引组(couplingphase),如以上组合一;把甲显性性状和乙隐性性状连系在一起遗传,而乙显性性状和甲隐性性状连系在一起遗传的杂交组合,称为相斥相或相斥组(repulsionphase),如以上组合二。(二)连锁遗传的解释1.果蝇眼色与翅长连锁遗传:相引相2.果蝇眼色与翅长连锁遗传:相斥相3.结果:F1形成四种类型的配子;但比例显然不符合1:1:1:1,且亲本类型配子明显多于重组型配子;两种亲本型配子数大致相等,两种重组型配子数也大致相等。4.连锁遗传现象的解释:(三)完全连锁和不完全连锁完全连锁(completelinkage)灰身(b+)对黑身(b)为显性,长翅(vg+)对残翅(vg)为显性不完全连锁(incompletelinkage)二、交换以相引组为例,其F1为:可见,两对连锁基因之间发生交换的孢母细胞的百分数,恰恰是重组型配子(又称交换型配子)百分数的2倍。一、交换值的概念二、交换值的测定由图5-7的测交结果可以求得:重组型配子数=149+152=301总配子数=4032+149+152+4035=8368交换值=×100=3.6%(二)、自交法小麦、水稻、豌豆及其它自花授粉作物不仅去雄和授粉比较困难,而且进行一次授粉只能得到少量种子。因此,一般利用自交法测定交换值。自交法的原理与过程:(以香豌豆花色与花粉粒形状两对相对性状,P-L交换值测定为例。)香豌豆P-L基因间交换值测定:2.F2的4种表现型(9种基因型)及其理论比例为:P_L_(PPLL,PPLl,PpLL,PpLl):a2+2ab+2ac+2bc+2adP_ll(PPll,Ppll):b2+2bdppL_(ppLL,ppLl):c2+2cdppll:d23.而F2中双隐性个体(ppll)的实际数目是可出直接观测得到的(本例中为1338),其比例也可出直接计算得到(1338/6952),因此有:三、遗传距离四、影响交换值的因素一、基因定位(genelocation/localization)*基因定位的三个层次:染色体定位(单体、缺体、三体定位法);染色体臂定位(端体分析法);连锁分析(linkageanalysis)。(一)、两点测验(two-pointtestcross)通过三次测验,获得三对基因两两间交换值、估计其遗传距离;每次测验两对基因间交换值;根据三个遗传距离推断三对基因间的排列次序。1.通过三次亲本间两两杂交,杂种F1与双隐性亲本测交,考察测交子代的类型与比例。例:玉米第9染色体上三对基因间连锁分析:子粒颜色:有色(C)对无色(c)为显性;饱满程度:饱满(SH)对凹陷(sh)为显性;淀粉粒:非糯性(Wx)对糯性(wx)为显性.(1).(CCShSh×ccshsh)F1×ccshsh(2).(wxwxShSh×WxWxshsh)F1×wxwxshsh(3).(wxwxCC×WxWxcc)F1×wxwxcc两点测验的3个测交结果2.计算三对基因两两间的交换值,估计基因间的遗传距离3.根据基因间的遗传距离确定基因间的排列次序并作连锁遗传图谱。C-Sh:3.6Wx-Sh:20两点测验的局限性(二)、三点测验(three-pointtestcross)一次测验就考虑三对基因的差异,从而通过一次测验获得三对基因间的距离并确定其排列次序。1.用三对性状差异的两个纯系作亲本进行杂交、测交,考察测交后代的表现型、进行分类统计。2.分析3对基因间的关系:3对均独立:8种表现型比例就应该彼此相等;2对连锁在一对同源染色体上,一对位于另一